The ventilation/perfusion (V/Q) ratio is an indication of how well alveolar ventilation matches pulmonary capillary perfusion.
Due to gravitational forces, the V/Q ratio ranges regionally in the lung, from 3.0 at the apex to 0.6 at the base
Clinicians and physiologists typically use the average value for the entire lung as a reference point.
Healthy average alveolar ventilation rate is approximately 4 liters of air/minute
Blood flow rate is approximately 5 liters of blood/minute.
A healthy V/Q for the entire lung is 0.8.
Healthy V/Q
Inspired air flows through the tracheobronchial tree and to the thin-walled, sac-like alveoli
Pulmonary capillaries are in close physical proximity
When the V/Q is 0.8, ventilation and perfusion are well matched and optimal gas exchange occurs (not perfect, which would be V/Q = 1; we’ll use the average for the entire lung).
The partial pressures of oxygen and carbon dioxide in the alveoli and pulmonary blood flow equilibrate (the specifics of gas exchange are discussed elsewhere).
V/Q mismatches, aka, defects, occur along a spectrum.
Two extremes of mismatches are shunts and dead space:
Shunts occur when the rate of alveolar ventilation is zero; V/Q = 0
Dead spaces occur where the rate of blood flow is zero; V/Q = infinity
Notice that the normal V/Q of 0.8 is between the two extremes (a V/Q of 1 would be perfect, but we are using the average value)
Mismatches between 0 and 0.8 reflect defects in alveolar airflow
Mismatches greater than 0.8 reflect defects in pulmonary capillary blood flow
Shunt
Most extreme alveolar ventilation defect.
Occurs when air flow is blocked and the alveoli remain unventilated.
Unventilated alveoli cannot participate in gas exchange with the pulmonary capillaries, which remain de-oxygenated.
As a result, the partial pressures of oxygen and carbon dioxide of the pulmonary blood remain equal to that of mixed venous blood.
Airway obstruction is a common cause of shunts
Because airflow ceases, alveolar collapse, aka, atelectasis, can occur in all or a portion of the lung.
Without proper ventilation, the partial pressure of arterial oxygen is reduced (aka, hypoxemia). In the case of shunts, hypoxemia cannot be reversed by administration of concentrated oxygen because oxygen does not reach the pulmonary blood flow.
Reduced ventilation
Alveoli are partially ventilated, so some gas exchange occurs with the nearby pulmonary capillary.
When ventilation is less than perfusion, the partial pressure of pulmonary blood carbon dioxide increases because it is held within the body, and the partial pressure of oxygen decreases because the normal amount of fresh oxygen is not available.
Reduced perfusion
Reduced perfusion causes an increased ventilation-to-perfusion ratio.
The partial pressure of pulmonary blood carbon dioxide decreases, while the partial pressure of oxygen increases.
Alveolar dead space
Absence of blood perfusion produces a ventilation-to-perfusion ratio equal to infinity.
No gas exchange occurs.
As a result, the partial pressures of oxygen and carbon dioxide of alveolar gas remain equal to that of inspired air.
Pulmonary embolism is a common cause of alveolar dead space
To minimize “wasted” ventilation, bronchiolar constriction diverts air from non-perfused alveoli.
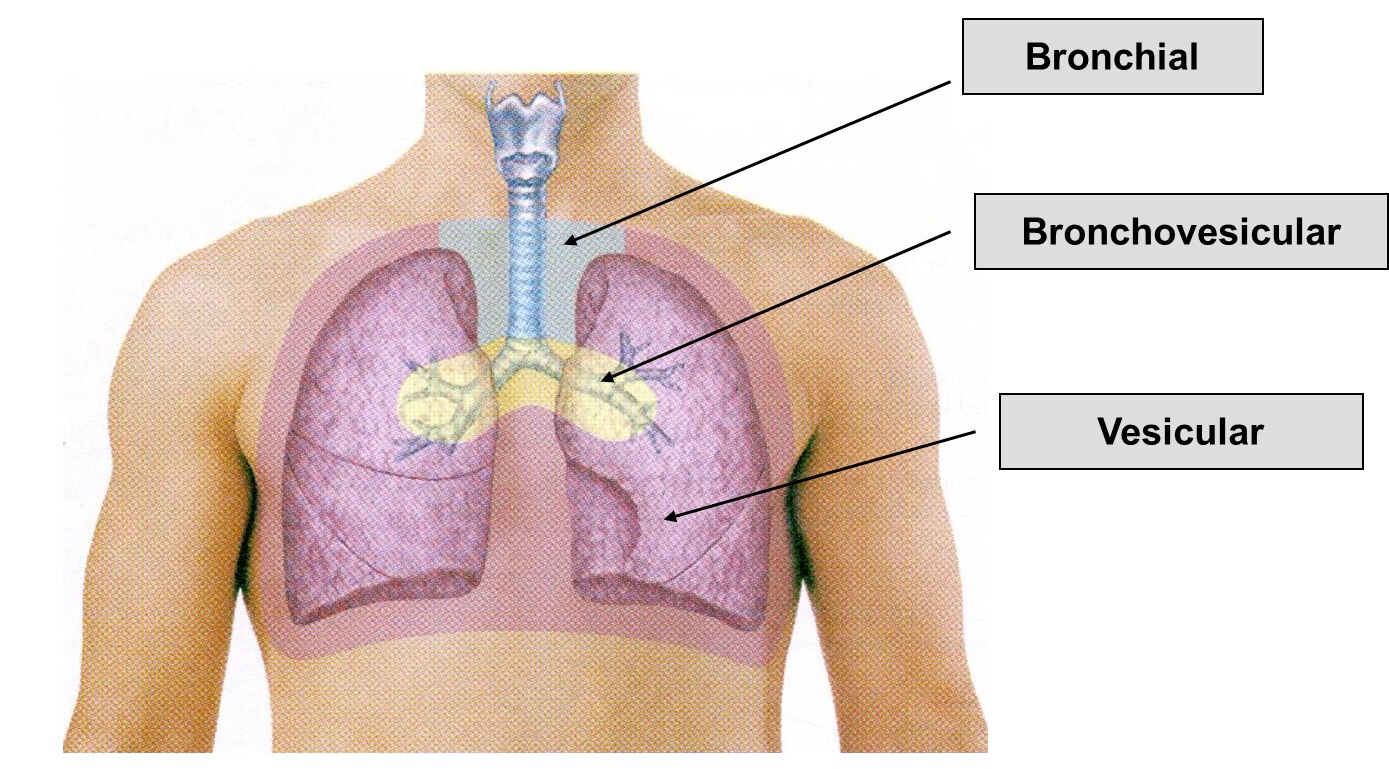
➡️ Vesicular breath sounds are heard across the lung surface. They are lower-pitched, rustling sounds with higher intensity during inspiration. During expiration, sound intensity can quickly fade. Inspiration is normally 2-3 times the length of expiration.
➡️ Various Types of Vesicular Breath Sound:
Exaggerated or puerile vesicular breath sounds: It is normal vescicular breathing with relatively greater clarity. It is common in children and thin built individual.
Diminished or absent breath sounds: Various causes are shallow breathing, airway obstruction, bulla, hyperinflation, pneumothorax, pleural effusion or thickening, and obesity.
Normal vesicular breath sound with prolonged expiration: It can occur in obstructive airway diseases like asthma and chronic bronchitis.
2) Bronchial breath sounds:
➡️ The bronchial breath sounds over the trachea has a higher pitch, louder, Expiratory phase is longer than inspiratory phase and there is a pause between inspiration and expiration.
➡️ It is normally heard anteriorly over the manubrium and posteriorly between the C7 and T3 vertebrae.
➡️ The vesicular breathing is heard over the thorax, lower pitched and softer than bronchial breathing.
Bronchial sound types👇🏻
3) Bronchovesicular breath sounds:
➡️ It is intermediate between bronchial and vescicular breathing. It has intermediate intensity and pitch with same duration of inspiratory and expiratory phase. It is normally heard anteriorly over 1st and 2nd intercostal spaces and between scapulae posteriorly.
Carbon dioxide is transported from the tissues to the blood, to the lungs, and out into the environment.
Three key forms of carbon dioxide present in the blood:
Dissolved CO2 constitutes about 5% of the total carbon dioxide content and contributes to the partial pressure; recall that the partial pressure of a gas is a major determinant of its diffusion.
Carbon dioxide bound to hemoglobin, constitutes about 3%, and is referred to as carbaminohemoglobin. The amount of carbaminohemoglobin in the blood is in part dependent upon the oxygen saturation:
The Haldane effect predicts that when blood oxygen increases, the affinity of hemoglobin for carbon dioxide decreases; in other words, when hemoglobin binds with oxygen, it more readily releases carbon dioxide.
Notice that this is the opposite of the Bohr effect, in which increased carbon dioxide reduces hemoglobin’s affinity for oxygen.
Bicarbonate is the chemically modified form of carbon dioxide that comprises the majority of carbon dioxide in the blood.
Be aware that the percentages of each type of carbon dioxide given here are approximations, because chemical reactions within the blood are ongoing.
Three sites of carbon dioxide:
Peripheral body tissues as byproduct of aerobic respiration.
Red blood cells in the vessels.
Lung.
Steps
Aerobic metabolism in the tissues produces carbon dioxide.
Partial pressure gradients between the tissues and the blood stream and also between the blood stream and the red blood cells, drives carbon dioxide diffusion into the red blood cells, where it mixes with H2O.
Within the red blood cell, show that carbonic anhydrase reversibly converts water and carbon dioxide to carbonic acid.
Carbonic acid dissociates to form hydrogen ions, which are buffered by hemoglobin, and bicarbonate, which can then exit the cell via chloride exchangers and enter the blood stream.
Bicarbonate enters the lungs, again via chloride exchangers; there, the reactions reverse to produce carbon dioxide, which is then expired during ventilation.
Hypercapnia
Occurs when ventilation is inhibited and carbon dioxide accumulates.
Hypercapnia can cause acidosis (low blood pH), which depresses the central nervous system.
This can lead to a suite of symptoms, including headache and confusion; if extreme, hypercapnia can lead to coma.
Oxygen delivery to the tissues is essential for life
It is dependent upon:
Cardiac output, which we’ve discussed in detail, elsewhere, and, the
Oxygen content of the blood, which will be the focus of this tutorial.
OXYGEN CONTENT
Amount of oxygen per unit volume of blood.
Amount of dissolved oxygen + hemoglobin-bound oxygen
Dissolved Oxygen
“Free” within the blood and can easily diffuse out of the vessel into the tissues (for example, recall that oxygen rapidly diffuses from the pulmonary capillaries to the alveoli in the lungs).
2% of the total oxygen content
Contributes to partial pressure, and therefore, drives diffusion.
Typically, the concentration of dissolved oxygen is ~ 0.3 mL of oxygen per 100 mL of blood
Oxygen Consumption:
Given that the average rate of oxygen consumption in a person at rest is ~ 250 mL of oxygen per minute, the tissues cannot rely on dissolved oxygen, alone. Thus, additional oxygen must be held within the body, but also readily available to the tissues.
Hemoglobin solves this problem by reversibly binding oxygen and delivering it in the bloodstream to the tissues.
Hb-Bound O2
Hemoglobin is a globular protein, which comprises four subunits, each of which can bind a single oxygen molecule.
Can bind up to four total oxygen molecules, but can be bound to fewer.
Hemoglobin-bound oxygen comprises 98% of the total oxygen content; thus, it is a major contributor to total oxygen content and delivery.
Amount of Hb is determined by two variables:
The percentage of saturation
The oxygen-binding capacity of hemoglobin present in the blood
Saturation percentage
Refers to the percentage of hemoglobin subunits bound to oxygen; hemoglobin bound to two oxygen molecules is 50% saturated, and Hb bound to four oxygen molecules is 100% saturated.
Oxygen partial pressure determines the saturation percentage
The oxyhemoglobin dissociation curve illustrates this relationship.
Sigmoid curve demonstrates how hemoglobin saturation changes in response to increasing partial pressure of oxygen.
Steep portion of the curve is due to positive cooperative binding: each time hemoglobin binds an oxygen molecule, its affinity for oxygen increases. It’s as if hemoglobin is offered potato chips; after it gets one, it “craves” more.
Healthy systemic arterial blood is nearly 100% saturated
Additional information about the oxyhemoglobin dissociation curve
“P 50” reflects the partial pressure value at which hemoglobin reaches 50% saturation.
If the curve shifts left or right, the P 50 will change to reflect hemoglobin’s altered affinity for oxygen.
These changes can be predicted, as follows:
Factors that shift the curve to the right decrease hemoglobin’s affinity for oxygen, and increase the P50 value; in other words, hemoglobin readily releases oxygen at lower partial pressures.
Factors that cause a leftward shift have the opposite effects: affinity is increased, and the P50 value decreases.
Some common causes of shifts include:
Increases in carbon dioxide and subsequent decreases in pH are shift the curve to the right; this phenomenon, called the Bohr effect, ensures that oxygen delivery meets tissue demand.
Alternatively, a decrease in carbon dioxide and increase in pH will increase affinity; this conserves oxygen when demand is low.
Increased body temperature, such as during strenuous activity, oxygen release is made easier, and, vice versa.
Increased altitude induces hypoxia, which decreases hemoglobin’s affinity for oxygen to ensure oxygen release to the tissues, and,
Fetal hemoglobin (Hemoglobin F) causes a leftward shift; increased affinity facilitates oxygen loading from the maternal blood supply, despite very low placental partial pressure oxygen levels.
Oxygen-binding capacity
Second variable needed to calculate the amount of oxygen-bound hemoglobin.
Oxygen-binding capacity is the maximum amount of oxygen bound to hemoglobin at 100% saturation.
It depends upon two variables: hemoglobin concentration and hemoglobin’s capacity to bind oxygen.
Standard values
Hemoglobin concentration is 15 g/100 mL
1 gram of hemoglobin A, the adult form, can bind 1.34 mL of oxygen.
So, typical oxygen-binding capacity: 15 grams/100 mL * 1.34 mL oxygen = 20.1 mL/oxygen per 100 mL blood;
Now, we can say that with oxygen saturation at 100%, the total amount of oxygen-bound hemoglobin is 20.1 mL of oxygen per 100 mL of blood.
Solve equation for total oxygen content:
Amount of dissolved oxygen equals 0.3 mL oxygen/100 milliliters of blood
Amount of hemoglobin-bound oxygen is 20.1 mL oxygen/100 milliliters of blood
Total oxygen content of blood is 20.4 mL oxygen per 100 milliliters of blood
Keep in mind that the values given here are for reference; physiological and pathological variations will alter the total oxygen content of blood, and, therefore, its delivery to the tissues.
Gas transport across the alveolar and pulmonary capillary walls is either perfusion limited or diffusion limited.
The partial pressure gradient of oxygen between the alveoli and the pulmonary blood flow drives diffusion; this is true whether transport is perfusion or diffusion limited.
Oxygen transport is perfusion-limited in healthy lungs.
Diffusion-limited in fibrosis (a pathology) and high altitude (an environmental change).
PERFUSION-LIMITED TRANSPORT OF OXYGEN
When the partial pressure gradient across the alveolar-capillary wall drops to zero, net diffusion ceases.
Only way to increase transport is to increase in capillary blood perfusion increases transport, which is another way of saying that transport is limited by perfusion.
Graph illustrates:
Partial pressure of oxygen of inspired alveolar gas is constant, at 100 mmHg.
Mixed venous blood arrives at the pulmonary capillary with a partial pressure of 40 mmHg.
As it passes through the capillary, its partial pressure of oxygen rapidly increases.
At approximately 1/3rd of the way through the capillary it equilibrates with that of the alveolar gas.
No further net diffusion occurs.
The area between the alveolar and arterial oxygen partial pressures represents the decreasing gradient between the two.
DIFFUSION-LIMITED TRANSPORT:
The partial pressure gradient is maintained; that is, equilibrium between alveolar and pulmonary oxygen is not reached before the pulmonary blood leaves the capillaries.
Example 1:
Pulmonary fibrosis, a disease in which the alveolar walls are thickened.
As the distance for diffusion increases (through lung fibrosis), the rate of oxygen transport slows, and, consequently, total oxygen transfer is reduced, which leads to hypoxemia (low partial pressure of oxygen in the blood).
Graph illustrates:
Partial pressure of oxygen in the alveolar gas remains constant, at 100 mmHg.
Partial pressure of oxygen in the mixed venous blood begins at 40 mmHg, but, because diffusion is impaired and its rate decreased, never reaches equilibrium with the alveoli.
Shaded area that represents the partial pressure gradient between the alveoli and systemic arterial blood remains wide along the length of the capillary.
As we can see, it is the slower rate of diffusion that limits the total transport of oxygen from the alveoli to the pulmonary blood.
Example 2:
High altitude.
Drop in atmospheric partial pressure of oxygen leads to a drop in the partial pressure of oxygen in the alveoli.
To compensate, hemoglobin’s affinity for oxygen is reduced, so that oxygen is more easily released to the body tissues; this shifts the oxyhemoglobin curve to the right (discussed in detail elsewhere) to increase diffusion.
Gas exchange in the lungs involves the diffusion of oxygen and carbon dioxide between the lungs and peripheral tissues.
Partial pressure gradient is a key driver of diffusion;
In healthy lungs, oxygen and carbon dioxide diffuse rapidly and achieve equilibrium.
Dalton’s Law
Partial pressure of a gas is the pressure a gas would exert if it occupied the total volume of a mixture.
Example: Total pressure of the gases a mixture equals 7 mmHg; this is equal to the sum of: The partial pressure of “a,” which happens to be 4 mmHg, plus the partial pressure of “b,” which happens to be 3 mmHg.
Recognize that the partial pressure of a gas is not its concentration; however, Henry’ Law states that the concentration of a gas is dependent, in part, upon its partial pressure.
PARTIAL PRESSURE DRIVES DIFFUSION
Key Points:
Dry inspired air (P I)
Oxygen partial pressure = 160 mmHg
Carbon dioxide partial pressure = 0 mmHg.
As it moves through the moist trachea, the oxygen is “diluted” by water vapor, so,
Humidified tracheal air partial pressures:
Oxygen ~150 mm Hg
Carbon dioxide remains unchanged = 0 mmHg.
Partial pressures of oxygen and carbon dioxide within the mixed venous blood of the capillary:
Partial pressure of oxygen is ~ 40 mmHg; this relatively low value reflects the metabolic activity of the peripheral tissues, which have removed much of the oxygen from the blood.
Partial pressure of carbon dioxide is ~ 46 mmHg; this relatively high value reflects the production of carbon dioxide by the peripheral tissues.
As the pulmonary blood passes by the alveolus, gas exchange occurs and rapidly reaches equilibrium so that:
Oxygen diffuses from the alveolus to the capillary, so that both alveolar and systemic arterial blood partial pressure is 100 mmHg, and,
Carbon dioxide diffuses from the capillary to the alveolus, which increases the alveolar partial pressure of carbon dioxide to 40 mmHg (from 0), and, reduces the systemic arterial blood partial pressure of carbon dioxide to 40 mmHg.
Perfusion-limited Diffusion:
Notice that, because diffusion occurs rapidly and equilibrium is achieved, the partial pressure gradient is negated
The only way to increase gas exchange at this point would be to increase pulmonary blood perfusion and the rate at which mixed venous blood arrived at the alveoli.
Rate of diffusion of oxygen and carbon dioxide
Fick’s Law:
Vgas = (D*(P1-P2)*A)/ T
D = Diffusion coefficient (aka, constant) specific to that gas (which depends upon the solubility of the gas and the square root of the molecular weight) P1-P2 = The partial pressure gradient, the driving force of diffusion A = Surface area available for diffusion T = Thickness of the membrane barrier
Clinical Correlation:
With the diffusion rate equation, we can predict that respiratory diseases will negatively affect diffusion rates:
Emphysema is characterized by alveolar destruction, and, therefore, decreased surface area available for diffusion.
Fibrosis causes thickening of the alveolar wall, and, therefore, increases the barrier to diffusion.
Functional Divisions of the Respiratory System: Conducting portion: the air passage conduit
Trachea
Bronchi
Bronchioles
Respiratory portion: the interface for gas (oxygen and carbon dioxide) exchange.
Respiratory bronchioles – Smooth muscle in respiratory bronchiole wall contracts to control airflow into alveolar sacs.
Alveolar ducts: connect bronchioles to thin-walled alveolar sacs.
Alveoli: grape-like sacs with a large surface area for gas exchange. – Dense capillary network on alveolar sac maximizes surface area and gas exchange efficiency.
Alveolar sac
Functional unit of the lung
Specialized alveolar cells: – Type I cells:simple squamous epithelial cells that form alveolar walls → very thin, allowing easy oxygen and carbon dioxide diffusion; most abundant. – Type II cells: secrete surfactant. – Alveolar macrophages: clear debris and perform immune functions; circulate within alveolar lumen. – Interstitial fluid bathes the alveolus and capillaries that surround it. – Elastin fibers in interstitial fluid assist in pulmonary elastic recoil and efficient expiration of air.
Surface Tension in Lungs & Surfactant
Water produces surface tension via its cohesive properties (hydrogen bonding).
Surfactant = lipid- and protein-rich fluid secreted from Type II cells. – Covers Type I cells. – Reduces surface tension. – Prevents alveolar collapse upon expiration. – Without surfactant, surface tensions causes alveoli collapse.
Premature infants are unable to produce surfactant.
Causes increased alveolar surface tension.
Alveoli collapse upon expiration.
Treated with surfactant replacement until their Type II cells produce surfactant, themselves.
The Basics of Gas Exchange
Gas exchange occurs across the respiratory portion of the lungs via diffusion.
Diffusion = movement of solutes (CO2 and O2) along their concentration gradient, from areas of high concentration to areas of low concentration.
Circulation: – The heart delivers deoxygenated blood to the lung capillary via the pulmonary arteries. The pulmonary arteries are the only arteries that carry deoxygenated blood, and the lungs are the only tissue to receive deoxygenated blood. – Lung capillaries return oxygenated blood to the heart via the pulmonary veins. – Concentration gradients differ in the lungs and the peripheral tissues → determine direction of diffusion.
Gas Exchange in the Alveoli
Concentration gradients:
Alveolar lumen: high O2, low CO2 concentration. – We breathe in oxygen-rich air!
Alveolar capillary: low O2, high CO2 concentration. – Most of blood oxygen has been used in peripheral tissue. – Blood accumulates metabolic wastes (CO2) as it circulates.
Creates concentration gradient → allows diffusion of O2 and CO2
Diffusion of O2 and CO2 down their concentration gradients:
O2 diffuses from the alveolar lumen → across the interstitial space → into the capillary, picked up by hemoglobin.
CO2 diffuses from capillary → across interstitial space → into alveolar lumen.
Gas Exchange in the Peripheral Tissues
The systemic circuit:
Heart delivers O2-rich blood to a peripheral capillary via the aorta.
Peripheral tissues return deoxygenated blood to the heart via the venae cava, which concludes the systemic circuit.
CO2 and O2 diffuse down their gradients just like in the lungs.
Concentration gradients:
Peripheral Tissue: low O2, high CO2. – CO2 accumulates as a metabolic byproduct. – Peripheral Capillary: high O2, low CO2.
Creates concentration gradient → allows diffusion of O2 and CO2
Diffusion of O2 and CO2 down their concentration gradients:
O2 diffuses from capillary →peripheral tissues.
CO2 diffuses from the peripheral tissues → capillary.
Blood returns to the lungs → CO2 diffuses out of the blood → CO2 exhaled.
Summary:
Oxygen diffuses into alveolar capillaries and out of the peripheral capillaries.
Carbon dioxide diffuses out of alveolar capillaries and into the peripheral capillaries.
Respiratory dead space and its relation to the alveolar partial pressure of oxygen.
Key points:
Gas exchange requires the close physical association of ventilated alveoli and perfused pulmonary capillaries.
However, in the dead spaces of the respiratory tract, one (or both) of these requirements is absent, and gas exchange does not occur.
Key points:
Gas exchange requires the close physical association of ventilated alveoli and perfused pulmonary capillaries.
However, in the dead spaces of the respiratory tract, one (or both) of these requirements is absent, and gas exchange does not occur. Anatomic dead space: conduction portion of the respiratory tract (we show the tracheobronchial tree in this image). Functional dead space: aka, alveolar dead space comprises alveoli where gas exchange does not occur (i.e., non-perfused alveoli). Physiologic dead space: includes the anatomical space and functional dead space; this is the total volume of the respiratory tract that does not participate in gas exchange. It can be calculated using the Bohr equation.
Ventilation
Minute ventilation rate
The total rate of air-flow into and out of the lungs.
Includes the air-flow through the tracheobronchial tree and to both the functional alveoli and non-functional alveoli.
Alveolar ventilation rate
Refers to the rate of air-flow into and out of the functioning alveoli, only (not though the physiologic dead spaces).
Partial Pressure of Alveolar O2
Minute and alveolar ventilation rate can be used to calculate the partial pressure of alveolar oxygen, which is a key facet of clinical respiratory evaluations.
Calculation Steps: – Calculate the minute ventilation rate: VE = TD x Breaths/Min Recall that tidal volume can be measured clinically using a spirometer. – VA = VE – VD (clinically determined via the Bohr equation). – Alveolar ventilation equation states that: PACO2 = (VCO2 x K)/VA K is usually 863 mmHg; this constant controls for variation in physical measurement conditions. – Alveolar gas equation states: PAO2 = PIO2 – (PACO2/RQ) RQ = respiratory quotient (aka, respiratory exchange ratio) is typically 0.8; it describes the ratio of the amount carbon dioxide produced in metabolism to the amount of oxygen consumed.
Summary
When alveolar ventilation rate decreases, the alveolar partial pressure of carbon dioxide increases while the partial pressure of oxygen decreases.
This makes intuitive sense, as you can imagine the effects of reducing alveolar ventilation by holding your breath:
Carbon dioxide is held in your lungs, and the amount of oxygen will necessarily decrease because you aren’t bringing fresh air into your lungs.
More specifically, the alveolar gas equation predicts the degree of change in alveolar partial pressure of oxygen for a given change in the partial pressure of alveolar carbon dioxide, as long as the respiration quotient stays at 0.8: For example, if alveolar ventilation is reduced by one half, alveolar partial pressure of carbon dioxide increases two-fold, and partial pressure of oxygen decreases by slightly more than one-half.
Notice that the alveolar gas equation also informs us that if the respiratory quotient changes, the relationship between alveolar partial pressure of carbon dioxide and oxygen will also change.
Clinical Correlation:
In healthy lungs, the the alveolar and systemic arterial partial pressure oxygen gradient is low, between 5-15 mmHg, because oxygen freely diffuses across the alveolar wall into the pulmonary capillaries.
However, an oxygen gradient above 30 can indicate pulmonary malfunction because oxygen transport is significantly impaired.
Release carbon dioxide: rids body of metabolic waste.
Regulate blood pH: via regulation of blood oxygen and carbon dioxide concentrations.
Three major components:
Muscular and skeletal components of the chest wall (“the pump”).
Alveoli = interfaces for gas exchange (“gas exchanger”).
Areas of the brain that stimulate respiratory function (“the controller”).
Boyle’s Law and The Respiratory System:
Boyle’s law describes the relationship between the pressure and volume of a gas → dictates the mechanics of respiration.
Boyle’s Law Equation: (P1)(V1) = (P2)(V2)
Pressure and volume of a gas are inversely related (assume temperature is constant).
Key anatomical/mechanical structures
Pair of lungs connected by a trachea
Lungs sit within a double-walled pleural sac, which separates lungs from thoracic wall.
Pleural space, which is the potential space within sac.
Sternum and ribs around pleural sac
The internal and external intercostal muscles lie between the ribs
Diaphragm comprises a sheet of skeletal muscle and forms bottom border of thoracic cavity. – Contraction/relaxation changes the volume of the thoracic cavity, which alters the intrapulmonary and intrapleural pressures in accordance with Boyle’s law.
Air pressures
Dictate movement of air during inspiration and expiration
Atmospheric pressure = 760 mm Hg, external to the thoracic cavity.
Intrapulmonary pressure (intra-alveolar pressure) = atmospheric pressure at rest, refers to the pressure within the lungs
Intrapleural pressure = 756 mmHg, pressure in the pleural space.
Inspiration: actively increases thoracic volume.
Movements during Inspiration:
Diaphragmatic Contraction:
Diaphragm flattens and moves inferiorly as it contracts.
External Intercostal Muscle Contraction:
External intercostal muscles contract
Forces sternum to move in an anterior and superior direction
Effects:
As thoracic volume increase: – Intrapulmonary pressure decreases to below atmospheric pressure; creates pressure gradient that facilitates inhalation. – Air moves down this pressure gradient: Moves from atmosphere (high P) through trachea into lungs (low P)
Gas exchange occurs in lungs → need to exhale carbon dioxide
Expiration passively decreases thoracic volume.
Movements during Expiration: – Diaphragm relaxes – Ribs back to resting position (move medially) – External intercostal muscles relax – Sternum back to resting position (moves inferiorly)
Effects:
As thoracic muscles relax: – Lungs elastically recoil to their pre-inspiratory volume; Lung volume decreases. – Intrapulmonary pressure increases above atmospheric pressure; creates pressure gradient that facilitates expiration. – Air moves down pressure gradient: Moves from lungs → out trachea → into atmosphere, CO2 exhaled.